2015年2月12日,中國科學院北京基因組研究所楊運桂研究組與中國科學院動物研究所周琪研究組及中國科學院遺傳與發育生物學研究所王秀杰研究組,整合三方在RNA m6A甲基化、干細胞和生物信息的研究優勢,合作開展“RNA m6A甲基化的位點選擇性機制及其調控成體細胞重編程研究”,發現了RNA m6A甲基化位點選擇性機制及調控細胞重編程重要功能,研究成果以“m6A RNA methylation is regulated by microRNAs and promotes reprogramming to pluripotency”為題發表在Cell 子刊 Cell Stem Cell,并被選為16卷第3期封面特寫報道。
腺嘌呤第6位氮原子上的甲基化修飾 (N6-methyladenosine, m6A) 是高等生物mRNA中含量最為豐富的在進化上保守的修飾之一。m6A修飾具有序列特異性、甲基化位點選擇性及修飾水平動態性特征,由RNA m6A甲基轉移酶復合物WTAP/METTL3/METTL14催化形成及去甲基化酶ALKBH5和FTO催化去甲基化,并被結合蛋白YTHDF2和YTHDC1識別。m6A修飾調控靶基因mRNA的穩定性及選擇性剪切。但是m6A的分布特征、形成選擇性機制和在細胞命運決定中的作用在很大程度上還屬未知。
利用生化、干細胞、基因組學、生物信息學等多層次技術手段,合作團隊發現m6A修飾和非編碼microRNA在共同作用底物mRNA上的位點具有物理空間相關性:1-部分反向匹配潛在m6A修飾RRACH序列的非編碼microRNA能夠介導其結合的mRNA RRACH位點m6A甲基化;2-另一部分不反向匹配潛在m6A修飾RRACH序列的非編碼microRNA能夠介導結合的mRNA位點附近的RRACH序列被甲基化。在人和小鼠細胞中,過表達microRNA或其加工酶Dicer,可以顯著增加mRNA m6A修飾水平,而敲低兩者之一則導致m6A修飾水平下降。Dicer可以調控m6A甲基轉移酶催化亞基METTL3在細胞核內核小斑的定位。Dicer和microRNA均可以調控METTL3結合mRNA的親和力。過表達野生型microRNA可導致結合的mRNA甲基化位點m6A修飾水平升高,而突變體microRNA結合的新的mRNA位點原來為非甲基化位點則可以檢測到高m6A修飾水平。Dicer和microRNA的表達水平變化并不會影響m6A修飾的甲基轉移酶METTL3和去甲基化酶ALKBH5和FTO的蛋白表達水平,說明了非編碼microRNA能夠調控RNA m6A甲基轉移酶選擇底物mRNA的甲基化位點。
通過過表達RNA m6A甲基轉移酶催化亞基METTL3,可以顯著提高小鼠成纖維細胞轉化為多能性干細胞的重編程效率,而敲低METTL3則顯著降低小鼠成纖維細胞的重編程效率;并且使用抑制m6A形成的小分子抑制劑環亮氨酸處理和敲低METTL3效果一致,都可以顯著降低小鼠成纖維細胞的重編程效率;METTL3調控了細胞多能性因子Oct4、Sox2、Klf4和Myc的表達。
該項成果揭示了非編碼microRNA調控RNA m6A甲基轉移酶選擇底物mRNA的甲基化位點機制和RNA m6A修飾調控細胞重編程重要功能。為進一步研究m6A的生物功能和RNA表觀遺傳提供了依據,為研究與正常生理(如干細胞干性維持和分化)或異常病理生命活動(如惡性腫瘤等)關聯分子機理提供了新的表觀調控研究方向。
該研究得到科技部重大研究計劃、中科院戰略性先導專項、國家自然科學基金等項目的資助。
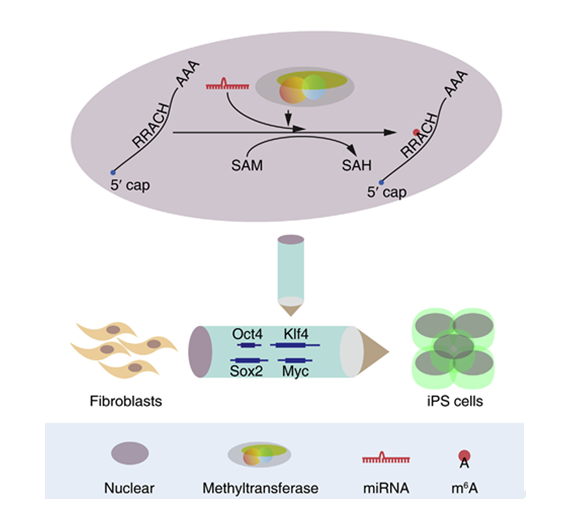
RNA m6A甲基化位點選擇性機制及其調控細胞重編程重要功能
論文鏈接


